Křeček syrský
Genetika
Základy
"Po většinu času byly naše geny záhadou.
Pouze v poslední době byl genetický kód rozluštěn, mnoho genů lokalizováno
a mnohým genům porozuměno. Naneštěstí křečci syrští nepatřili k
vybraným druhům, které byly detailně prozkoumány. Protože jejich geny
nemohou být takto přesně lokalizovány, musíme pouze odvozovat to, co je nám
skryto z toho, co vidíme, když se díváme na zvíře. Dokonce i přes pečlivé
pozorování se dá vydedukovat jen část genetických informací."
Použito
se svolením Jan,
The River Road
Hamstery http://hometown.aol.com/theriverrd/genetics.htm
Není až zase tak těžké porozumět genetice, je však
nutné si uvědomit, že vše souvisí se vším. Začnu tedy od úplného začátku
a pokusím se vše trošku zjednodušit. Pro chovatele je znalost základů
genetiky nezbytně nutná pro rozhodování při křížení, aby se předešlo
určitým chybám a omylům.
Tedy křeček syrský se skládá z různých částí -
orgánů, kostí atd., které jsou všechny složeny z buněk. Buňka je základní
stavební jednotka organismů. Skládá se z jádra, jadérka, jaderné membrány,
endoplazmatického retikula, mitochondrií, cytoplazmy a cytoplazmatické membrány
(samozřejmě mimo jiné). Půjdeme - li ještě více do hloubky, dostaneme se
až na makromolekulární úroveň (bílkoviny, nukleové kyseliny), a dále k
jednotlivým atomům chemických prvků, protonům, elektronům a neutronům, až třeba na úroveň nejmenších
zjištěných částí atomů - kvarkům a partonům. Nás bude ovšem nejvíce
zajímat jádro buňky, chromozómy v něm uložené a DNA.
Genetika je nauka o dědičnosti a proměnlivosti živých
soustav. Je vědou poměrně mladou, za zakladatele je považován Gregor Johann
Mendel, který se narodil a žil na území dnešní České republiky (ve
Slezsku a na Moravě), ovšem sám se považoval za Němce, a jeho rodnou řečí
byla němčina. Zabýval se hybridizačními pokusy u rostlin a roku 1866 vydal
o svých pozorováních práci nazvanou Versuche über Pflanzenhybriden (Pokusy
s rostlinnými kříženci), ve které jsou popsány v budoucnu nazvané
Mendelovy zákony. Ve své době však neměla jeho práce vůbec žádný ohlas
a byla dokonce zapomenuta. Ke znovuobjevení Mendelovy práce a ke vzniku
genetiky jako plnohodnotného vědního oboru tak dochází až na počátku 20.
století. Klíčovým okamžikem byl samozřejmě objev DNA - nositelky genetické
informace, která byla poprvé prokázána již v roce 1944 týmem Američana
Oswalda T. Aweryho, další poznatky ohledně komplementarity bází přinesl
Erwin Chargaff a na jejich práci navazují James D. Watson a Francis H. Crick,
kteří roku 1953 předložili strukturní model dvoušroubovice DNA. V roce
1956 je stanoven počet chromozómů v lidské buňce, v roce 1963 byl rozluštěn
genetický kód (způsob, jakým je informace zakódovaná v DNA čtena a překládána
do jazyka bílkovin). Objev moderních sekvenovacích principů umožnil
sekvenování genomů jednoduchých organismů (1965 - genom kvasinky), s rozvíjejícím
se technickým pokrokem bylo možné sekvenovat stále větší genomy, což
vyvrcholilo sekvenováním lidského genomu vybraných částí roku 2001 a
kompletní sekvence lidského genomu spatřila světlo světa až v dubnu roku
2003. Nových objevů se v oblasti genetiky budeme tedy dočkávat každým
dnem...
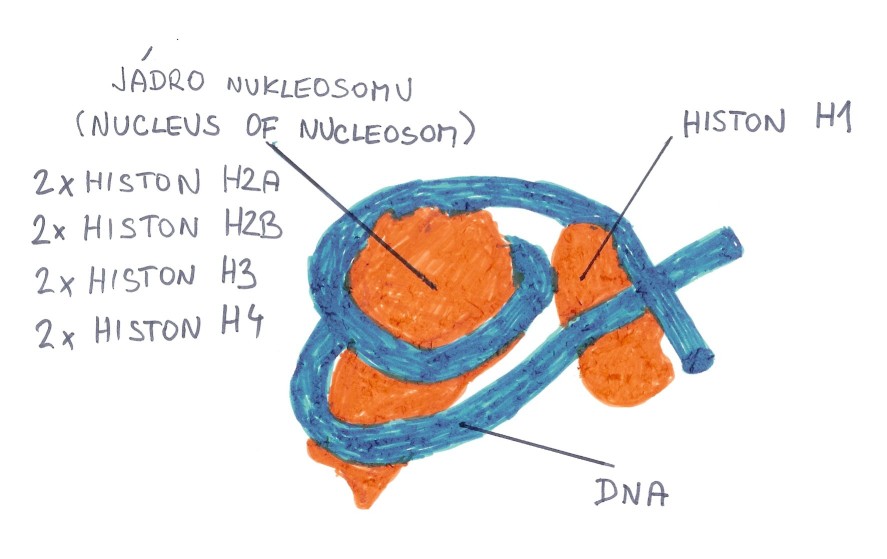 V nitru buňky se nachází jádro a v něm chromozómy.
Chromozómy jsou pentlicovité útvary, jejichž základní stavební jednotkou
jsou tzv. nukleosomy - což je útvar tvořený 8 histony obtočený DNA.
Histony jsou malé bazické bílkoviny, které mají vysoký obsah kladně nabitých
aminokyselin (lysin a arginin). Pomocí tohoto kladného náboje vytvářejí
reverzibilní komplexy s DNA. Rozlišuje se pět typů
histonů: H1, H2A, H2B, H3 a H4. Histony H2A, H2B, H3 a H4 tvoří vždy ve dvou
kopiích oktamery - jádro nukleosomu, kolem něhož se obtáčí dvojšroubovicová
DNA. Histon H1 je přítomen v menším množství než ostatní histony, a ačkoli
je též vázán na DNA, není součástí nukleosomů.
Spiralizací nukleosomů vznikají chromatinová vlákna a další spiralizací
těchto vláken vznikají již celé chromozómy.
V nitru buňky se nachází jádro a v něm chromozómy.
Chromozómy jsou pentlicovité útvary, jejichž základní stavební jednotkou
jsou tzv. nukleosomy - což je útvar tvořený 8 histony obtočený DNA.
Histony jsou malé bazické bílkoviny, které mají vysoký obsah kladně nabitých
aminokyselin (lysin a arginin). Pomocí tohoto kladného náboje vytvářejí
reverzibilní komplexy s DNA. Rozlišuje se pět typů
histonů: H1, H2A, H2B, H3 a H4. Histony H2A, H2B, H3 a H4 tvoří vždy ve dvou
kopiích oktamery - jádro nukleosomu, kolem něhož se obtáčí dvojšroubovicová
DNA. Histon H1 je přítomen v menším množství než ostatní histony, a ačkoli
je též vázán na DNA, není součástí nukleosomů.
Spiralizací nukleosomů vznikají chromatinová vlákna a další spiralizací
těchto vláken vznikají již celé chromozómy.
Molekula DNA je nositelkou dědičné informace, je velmi
dlouhá, pokud by se natáhla a změřila, dosáhla by délky až dvou metrů.
 Tato dlouhá molekula je tvořena dvojicí vláken stočených do pověstné
dvojité šroubovice. Ta vypadá jako zkroucený provazový žebřík, přičemž
roli "provazů" plní řetězec z molekul cukru a kyseliny fosforečné
a pomyslné "příčky" žebříku tu zastupují dvojice dusíkatých
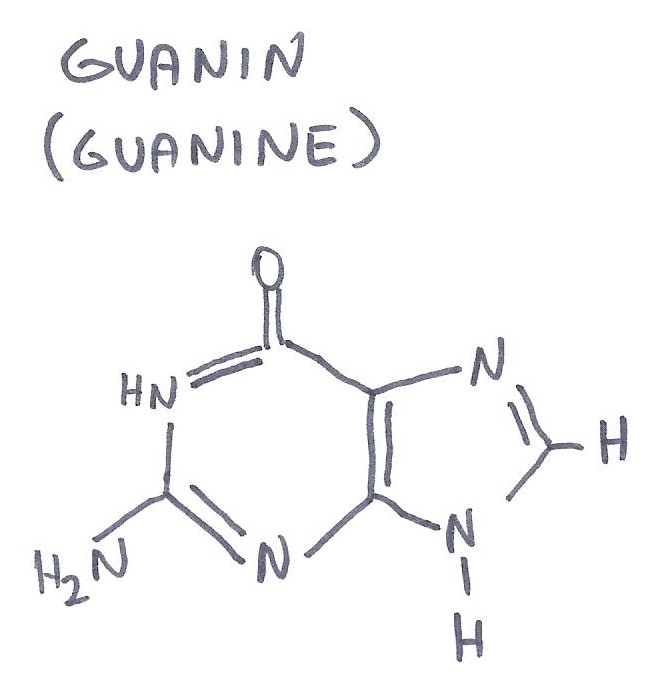
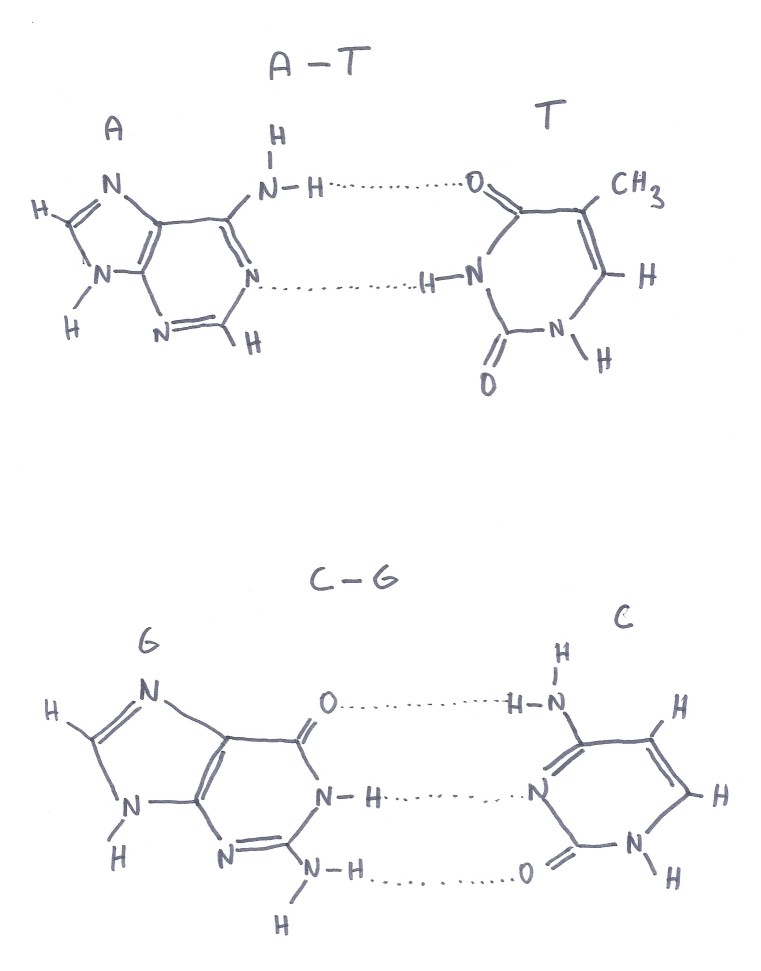
sloučenin tzv. bazí. Jedná se o čtyři typy bazí: adenin (A), guanin (G),
cytosin (C) a thymin (T), které se spojují vodíkovými můstky a to vždy jen
adenin s thyminem (dvěma vodíkovými můstky) a cytosin s guaninem (třemi vodíkovými
můstky). Různé molekuly DNA se liší sledem těchto dvojic bází a právě
tento kvartet A,C,G,T plní roli "písmen" genetického kódu, přičemž
důležité je jejich přesné pořadí. Navíc klíčové jsou relativně krátké
úseky čítající obvykle jen několik tisíc písmen genetického kódu. Těch
se např. v celé lidské dědičné informaci nachází asi 35 tisíc a zabírají
tu jen asi jen 1%. Pro tyto úseky DNA obvykle užíváme označení geny. Zbylých
téměř 99% dědičné informace se zdá být jen jakousi genetickou "vycpávkou",
ale to je zřejmě jen zdání, které klame. Možná, že nás ty nejpřekvapivější
objevy čekají právě v této "zbytečné DNA", která může sehrávat
klíčovou roli při řízení jednotlivých genů. Lidská a zvířecí (např.
křeččí) dědičná informace se velikostí ani počtem genů vzájemně téměř
neliší.
Tato dlouhá molekula je tvořena dvojicí vláken stočených do pověstné
dvojité šroubovice. Ta vypadá jako zkroucený provazový žebřík, přičemž
roli "provazů" plní řetězec z molekul cukru a kyseliny fosforečné
a pomyslné "příčky" žebříku tu zastupují dvojice dusíkatých
sloučenin tzv. bazí. Jedná se o čtyři typy bazí: adenin (A), guanin (G),
cytosin (C) a thymin (T), které se spojují vodíkovými můstky a to vždy jen
adenin s thyminem (dvěma vodíkovými můstky) a cytosin s guaninem (třemi vodíkovými
můstky). Různé molekuly DNA se liší sledem těchto dvojic bází a právě
tento kvartet A,C,G,T plní roli "písmen" genetického kódu, přičemž
důležité je jejich přesné pořadí. Navíc klíčové jsou relativně krátké
úseky čítající obvykle jen několik tisíc písmen genetického kódu. Těch
se např. v celé lidské dědičné informaci nachází asi 35 tisíc a zabírají
tu jen asi jen 1%. Pro tyto úseky DNA obvykle užíváme označení geny. Zbylých
téměř 99% dědičné informace se zdá být jen jakousi genetickou "vycpávkou",
ale to je zřejmě jen zdání, které klame. Možná, že nás ty nejpřekvapivější
objevy čekají právě v této "zbytečné DNA", která může sehrávat
klíčovou roli při řízení jednotlivých genů. Lidská a zvířecí (např.
křeččí) dědičná informace se velikostí ani počtem genů vzájemně téměř
neliší.
 |
|
 |
 DNA je zkratka pro kyselinu deoxyribonukleovou, což je
makromolekula tvořená řetězci vzájemně spojených deoxyribonukleotidů. Skládá
se z fosfátu, 5-ti uhlíkatého cukru deoxyribózy a dusíkaté báze (adenin,
guanin, cytosin či thymin). Její nejdůležitější schopností je schopnost
replikace. Při replikaci vznikají z původní
molekuly DNA dvě strukturně naprosto stejné dceřiné molekuly. Tímto způsobem
je zajištěna kontinuita genetických informací v následujících
generacích. Klíčovou roli při replikaci DNA mají enzymy (DNA polymerázy).
U člověka se vyskytuje 5 druhů enzymů označované jako DNA dependentní DNA
polymerázy. Při své práci vždy postupují od konce 5' ke konci 3'. Aby DNA
polymeráza mohla zahájit připojování nukleotidů nového vlákna DNA, musí
být vodíkové můstky - vazby mezi oběma vlákny - nejprve narušeny (využití
DNA dependentní RNA polymerázy). Místa, kde tato narušení vzniknou, jsou označována
jako replikační počátky. Vzhledem k tomu, že polymerázová aktivita je
pouze ve směru od 5' konce ke 3' konci, může tímto směrem probíhat
replikace pouze na jednom vlákně. Na tomto vláknu probíhá replikace kontinuálně
a označujeme je jako vedoucí řetězec. Na druhém řetězci je situace složitější.
Replikace zde probíhá proti směru rozplétání dvoušroubovice a to
diskontinuálně po menších úsecích. Tyto části se nazývají Okazakiho
fragmenty a celý řetěz nazýváme opožďující se řetězec. Takto vzniklé
fragmenty napojí k sobě do jednolitého vlákna enzym DNA ligáza. Posledním
enzymem, který je nezbytný pro replikaci, je DNA primáza. DNA polymeráza totiž
neumí zahájit polymeraci od jediného nukleotidu - proto zde nastupuje právě
DNA primáza (což je vlastně DNA dependentní RNA polymeráza), která
nasyntetizuje krátký úsek RNA - tzv. primer - od kterého už může DNA
polymeráza zahájit polymeraci. Takovýto primer vznikne nejen na vedoucím řetězci,
ale musí vzniknout i před každým Okazakiho fragmentem na opožďujícím se
řetězci. Primery jsou posléze vyštěpeny - chybějící úseky dosyntetizovány
a vlákno je spojeno DNA ligázou. DNA polymeráza udělá 1 chybu asi na 107
zreplikovaných bází.
DNA je zkratka pro kyselinu deoxyribonukleovou, což je
makromolekula tvořená řetězci vzájemně spojených deoxyribonukleotidů. Skládá
se z fosfátu, 5-ti uhlíkatého cukru deoxyribózy a dusíkaté báze (adenin,
guanin, cytosin či thymin). Její nejdůležitější schopností je schopnost
replikace. Při replikaci vznikají z původní
molekuly DNA dvě strukturně naprosto stejné dceřiné molekuly. Tímto způsobem
je zajištěna kontinuita genetických informací v následujících
generacích. Klíčovou roli při replikaci DNA mají enzymy (DNA polymerázy).
U člověka se vyskytuje 5 druhů enzymů označované jako DNA dependentní DNA
polymerázy. Při své práci vždy postupují od konce 5' ke konci 3'. Aby DNA
polymeráza mohla zahájit připojování nukleotidů nového vlákna DNA, musí
být vodíkové můstky - vazby mezi oběma vlákny - nejprve narušeny (využití
DNA dependentní RNA polymerázy). Místa, kde tato narušení vzniknou, jsou označována
jako replikační počátky. Vzhledem k tomu, že polymerázová aktivita je
pouze ve směru od 5' konce ke 3' konci, může tímto směrem probíhat
replikace pouze na jednom vlákně. Na tomto vláknu probíhá replikace kontinuálně
a označujeme je jako vedoucí řetězec. Na druhém řetězci je situace složitější.
Replikace zde probíhá proti směru rozplétání dvoušroubovice a to
diskontinuálně po menších úsecích. Tyto části se nazývají Okazakiho
fragmenty a celý řetěz nazýváme opožďující se řetězec. Takto vzniklé
fragmenty napojí k sobě do jednolitého vlákna enzym DNA ligáza. Posledním
enzymem, který je nezbytný pro replikaci, je DNA primáza. DNA polymeráza totiž
neumí zahájit polymeraci od jediného nukleotidu - proto zde nastupuje právě
DNA primáza (což je vlastně DNA dependentní RNA polymeráza), která
nasyntetizuje krátký úsek RNA - tzv. primer - od kterého už může DNA
polymeráza zahájit polymeraci. Takovýto primer vznikne nejen na vedoucím řetězci,
ale musí vzniknout i před každým Okazakiho fragmentem na opožďujícím se
řetězci. Primery jsou posléze vyštěpeny - chybějící úseky dosyntetizovány
a vlákno je spojeno DNA ligázou. DNA polymeráza udělá 1 chybu asi na 107
zreplikovaných bází.
Mutace jsou změny v
genotypu organismu oproti normálu. Většina mutací je náhodná, cílené
mutageneze se používají výhradně pro vědecké účely. V přírodě se udrží
pouze výhodné či neutrální mutace, protože nevýhodná mutace má za následek
většinou smrt mutanta či nevýhodné podmínky pro přežití. Příkladem by
mohla být barevná mutace křečka syrského stříbrná šedá, v takto barevné
srsti by zvíře brzy neuniklo pozornosti predátora, tudíž by nemělo šanci
dále předávat své zmutované geny. Do pouští a polopouští Sýrie, odkud
křeček syrský pochází, je nejvýhodnější zbarvení zlaté, které nejlépe
splyne s okolím. Mutace rozlišujeme podle místa a způsobu vzniku na genové
(probíhají na úrovni vlákna DNA - přidáním, ztrátou či náhradou bazí),
chromozómové (změní se struktura či počet chromozómů - znásobením, ztrátou,
převrácením části chromozómu, napojením části chromozómu na jiný
chromozóm či rozpadem chromozómu na fragmenty), genomové (znásobí se celé
chromozómové sady), dále mozaicismus (neoddělení chromozómů nastává až
během mitózy ve vyvíjejícím se organismu - na somatické úrovni) a
chimerismus (jedinec je tvořen dvěma liniemi buněk vzniklých ze dvou různých
zygot, které následovně splynuly v jednoho jedince - např. srostlá dvojčata).
Frekvence vzniku mutací je v čase přibližně konstantní a u příbuzných
druhů velmi podobná.
Mutageny jsou látky, které
jsou schopny vyvolat mutace. Dělí se na fyzikální (UV záření, RTG záření),
chemické (aromatické uhlovodíky - tabákový kouř, barviva, organická
rozpouštědla, hnojiva, herbicidy) a biologické (viry).
Gen je specificky uložená
jednotka dědičné informace. Z molekulárního hlediska jde o úsek nukleové
kyseliny se specifickým pořadím nukleotidů. Každý gen má své unikátní
místo na určitém chromozómu. Toto místo označujeme genový lokus. O
genech uložených na stejném chromozómu říkáme, že jsou spolu v genové
vazbě. Podle síly genové vazby se dědí tyto geny buď vždy společně
(silná vazba) nebo i odděleně (slabá vazba). Síla této vazby je nepřímo
úměrná vzdálenosti mezi geny - čím je větší vzdálenost, tím je slabší
vazba. Alela je konkrétní forma genu. V diploidní buňce existují pro
jeden gen vždy 2 alely. Alelu, která se vyskytuje běžně v populaci v přírodě,
nazýváme alelou divokou, neběžnou alelu alelou mutovanou. Jsou-li obě alely
stejné, jedinec je nazýván homozygotní, jsou-li různé heterozygotní.
Lokus však může mít i více alel - příkladem je lokus C.
Mezialelické vztahy stejného
genu jsou úplná dominance a recesivita, neúplná dominance a recesivita,
kodominance a superdominance. Dominantní alela potlačuje projev recesivní
alely a je zvykem ji označovat velkým písmenem. My chovatelé si vystačíme
pouze s dominancí a recesivitou bez označování, zda je úplná či neúplná,
a kodominancí. Jen pro vaši představu se úplná dominance projeví v plné síle
již u heterozygotů (páskovaný křeček syrský), neúplná až u homozygotů
(stříbrný šedý křeček syrský, kdy heterozygot Sgsg je "méně"
stříbrně šedý než homozygot SgSg). Pro naši definici tedy postačí, když
řekneme, že dominantní mutace je mutace, kdy stačí pouze jedna
mutovaná alela k projevu změny (páskovaný Ba_), recesivní mutace se
projeví pouze je-li přítomna na obou alelách (černý aa). Kodominance
je stav, kdy se obě přítomné alely u heterozygota projeví v celé míře a
navzájem se neovlivňují (heterozygot extrémně zředěný cdce).
Existuje však ještě genová interakce, při které se gen projeví jen při
současné přítomnosti jiného (druhého) genu, a polygenní dědičnost, při
které se na výsledném efektu podílí mnoho genů slabého účinku (barva očí,
výška).
Křeček syrský se rozmnožuje
pohlavně, s použitím pohlavních buněk (spermie, vajíčko), které se nazývají
gamety, přičemž samice je označována XX a samec XY. Pohlavní buňky nesou
polovinu dědičné informace nového jedince, tedy jen 22 chromozómů. Při
oplodnění dostává jedinec celou sadu 44 chromozómů, přičemž polovinu
genetické informace má od otce a polovinu od matky. Gamety vznikají v rodičích
redukčním dělením - meiosou - v praxi to znamená to, že rodič může
potomkovi předat pouze některé své geny - z každého genového páru pouze
jeden (jednu alelu). Zde bych ještě na okraj chtěla připomenout pravidla
Mendelovské dědičnosti - 3 Mendelovy zákony: 1. Mendelův zákon (o
uniformitě F1 - první generace potomků při křížení dvou homozygotů), 2.
Mendelův zákon (o náhodné segregaci genů do gamet - při křížení 2
heterozygotů může být potomkovi předána každá ze dvou alel (dominantní
i recesivní) se stejnou pravděpodobností) a 3. Mendelův zákon (o nezávislé
kombinovatelnosti alel).
Tady bych ukončila můj
dodatečný výklad toho, co jsem cítila jako potřebné doplnit k základům
genetiky, a dále budu již jen doslovně překládat vynikající práci Jan,
The River Road Hamstery.
Chromozómy a geny
"Téměř všechny
fyzické vlastnosti jsou zakódovány v molekulách DNA (na chromozómech), které
se nacházejí v jádrech většiny buněk těla. Chromozómy existují ve velmi
podobných párech, nazývaných homologické. Znaky zakódované v chromozómu
mají vždy svůj protějšek na sesterském chromozómu. Každá malá část
DNA obsahuje gen, a každý genový pár kontroluje přesně vzhled zvířete.
Daný pár genů (na sesterských chromozómech) může být úplně stejný
nebo velice jemně odlišný. Například v genovém páru, který řídí barvu
očí, jedna forma genu (která se nazývá alela) může kódovat pro
modrou, zatímco druhá pro hnědou. Mohou tam být další přídavné formy
genu pro barvu očí, ale jedinec může mít pouze dva z nich, které převládají
- po jednom na každém homologickém páru. Přesné místo genu v sadě
chromozómů se nazývá lokus a lokus je stejný pro všechny
jednotlivce daného druhu."
Dominantní a recesivní,
genotyp a fenotyp
"Každému genu, který
byl identifikován, se přidělí písmeno. Pokud alela změní vzhled zvířete,
když je přítomna pouze v jedné kopii, říká se, že je dominantní
a používá se pro ni velké písmeno. Recesivní alela musí být přítomna
ve dvou kopiích, aby se projevil její efekt, a označuje se malými písmeny.
Použijeme - li jako příklad
barvu očí, alela pro hnědé oči je dominantní a označuje se B. Alela pro
modré oči je recesivní a označuje se b. Protože jedinec musí mít dvě
alely pro každou vlastnost, možnosti genotypů (sestavy genetických kódů)
jsou BB, Bb a bb. Oba jedinci BB a Bb budou mít hnědé oči, protože hnědá
je dominantní a tudíž nemá možnost se projevit b v jedinci Bb. Pouze
jedinec s genotypem bb bude mít modré oči, protože nebude přítomna žádná
dominantní alela, která by převládla. Fenotyp je viditelná vlastnost
vyprodukovaná genotypem, anebo jinak, fenotyp je to, jak zvíře vypadá. Takže
BB a Bb jedinci mají sice rozdílný genotyp, ale úplně stejný fenotyp.
(Varovná poznámka, některé alely, přestože se označují velkými písmeny,
jsou neúplně dominantní, takže jedinec se dvěma dominantními
alelami bude mít odlišný vzhled od jedince s jednou dominantní a jednou
recesivní alelou.)"
Homozygotní a
heterozygotní
"Jestliže má jedinec
obě alely daného páru ve stejné formě, jako BB nebo bb, nazývá se homozygotní
pro tuto vlastnost. Pokud je forma odlišná, jako Bb, nazývá se heterozygot.
Homo - je předpona, která znamená stejný;
hetero - znamená různý."
Pohlavní chromozómy a pohlavní vazba
"Jen jeden speciální pár chromozómů u vyšších
živočichů nemá svůj přesný protějšek. Ve skutečnosti, nikdy nevypadají
stejně v mikroskopu. Jeden vypadá jako X a druhý jako Y, jedno rameno je
velmi zkrácené. Jejich tvary jim propůjčily jejich pojmenování, X a
Y. (Všechny ostatní chromozómové páry, kterým se říká autozómy,
jsou očíslované, ne pojmenované.) Živočich se dvěma X chromozómy je
samice; jedinec s jedním X a jedním Y je samec. Z
tohoto důvodu jsou X a Y nazývány pohlavní chromozómy. Je důležité
poznamenat, že geny na pohlavních chromozómech nejsou nezbytně vázány vždy
ve vztahu k pohlaví, nýbrž mohou určovat jakoukoliv vlastnost. Protože geny
na pohlavních chromozómech jsou přenášeny spolu s pohlavím, říká se jim
pohlavně vázané."
X-inaktivace
 "Na X chromozómu je mnoho genů, které jednoduše
nemají protějšek na Y chromozómu. Tyto geny u samců tedy určují jeho
fenotyp. U samic je však alela na obou X chromozómech jako u ostatních
chromozómů, nicméně to zde funguje trošku odlišně než u autozómů. Brzy
na počátku vývoje jedince je vybráno jedno X v každé buňce, které bude
aktivní po celý život samice, druhé X je odstaveno a zataraseno. Toto se nazývá
X-inaktivace, nebo někdy racionalizace či lyonizace. Každá z těchto prvotních
buněk, když se dále dělí a vytváří tkáň, předává přirozený sklon
k aktivaci stejného X. Nakonec je samice mozaiková, složená z některých
ploch pod nadvládou jednoho X a zbylých pod nadvládou druhého X."
"Na X chromozómu je mnoho genů, které jednoduše
nemají protějšek na Y chromozómu. Tyto geny u samců tedy určují jeho
fenotyp. U samic je však alela na obou X chromozómech jako u ostatních
chromozómů, nicméně to zde funguje trošku odlišně než u autozómů. Brzy
na počátku vývoje jedince je vybráno jedno X v každé buňce, které bude
aktivní po celý život samice, druhé X je odstaveno a zataraseno. Toto se nazývá
X-inaktivace, nebo někdy racionalizace či lyonizace. Každá z těchto prvotních
buněk, když se dále dělí a vytváří tkáň, předává přirozený sklon
k aktivaci stejného X. Nakonec je samice mozaiková, složená z některých
ploch pod nadvládou jednoho X a zbylých pod nadvládou druhého X."
Použito
se svolením Jan,
The River Road
Hamstery http://hometown.aol.com/theriverrd/basics.htm